Streptococcus pneumoniae (page 1)
(This chapter has 4 pages)
Introduction
Streptococcus pneumoniae is a normal inhabitant of the human upper respiratory tract. The bacterium can cause pneumonia, usually of the lobar type, paranasal sinusitis and otitis media, or meningitis, which is usually secondary to one of the former infections. It also causes osteomyelitis, septic arthritis, endocarditis, peritonitis, cellulitis and brain abscesses. Streptococcus pneumoniae is currently the leading cause of invasive bacterial disease in children and the elderly. Streptococcus pneumoniae is known in medical microbiology as the pneumococcus, referring to its morphology and its consistent involvement in pneumococcal pneumonia.
Pneumonia is a disease of the lung that is caused by a variety of bacteria including Streptococcus, Staphylococcus, Pseudomonas, Haemophilus, Chlamydia and Mycoplasma, several viruses, and certain fungi and protozoans. The disease may be divided into two forms, bronchial pneumonia and lobar pneumonia. Bronchial pneumonia is most prevalent in infants, young children and aged adults. It is caused by various bacteria, including Streptococcus pneumoniae.Bronchial pneumonia involves the alveoli contiguous to the larger bronchioles of the bronchial tree. Lobar pneumonia is more prone to occur in younger adults. A majority (more than 80%) of the cases of lobar pneumonia are caused byStreptococcus pneumoniae. Lobar pneumonia involves all of a single lobe of the lungs (although more than one lobe may be involved), wherein the entire area of involvement tends to become a consolidated mass, in contrast to the spongy texture of normal lung tissue.
Bacteriology
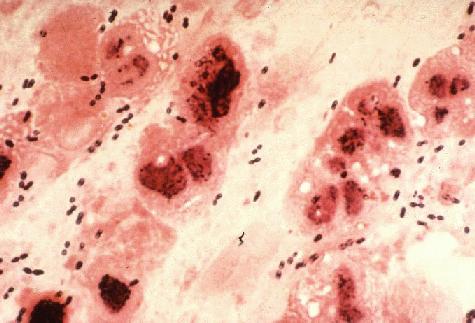
Gram Stain of a film of sputum from a case of lobar pneumonia. CDC.
Streptococcus pneumoniae Gram-stain of blood broth culture. CDC.
Streptococcus pneumoniae A mucoid strain on blood agar showing alpha hemolysis (green zone surrounding colonies). Note the zone of inhibition around a filter paper disc impregnated with optochin. Viridans streptococci are not inhibited by optochin.
Serotyping

Streptococcus pneumoniae Quellung (capsular swelling) reaction can be used to demonstrate the presence of a specific capsular type of the bacterium.

Streptococcus pneumoniae scanning electron micrograph of a pair of diplococci. CDC.
Streptococcus pneumoniae Fluorescent antibody stain of capsular material. CDC.
(This chapter has 4 pages)
Introduction
Streptococcus pneumoniae is a normal inhabitant of the human upper respiratory tract. The bacterium can cause pneumonia, usually of the lobar type, paranasal sinusitis and otitis media, or meningitis, which is usually secondary to one of the former infections. It also causes osteomyelitis, septic arthritis, endocarditis, peritonitis, cellulitis and brain abscesses. Streptococcus pneumoniae is currently the leading cause of invasive bacterial disease in children and the elderly. Streptococcus pneumoniae is known in medical microbiology as the pneumococcus, referring to its morphology and its consistent involvement in pneumococcal pneumonia.
Pneumonia is a disease of the lung that is caused by a variety of bacteria including Streptococcus, Staphylococcus, Pseudomonas, Haemophilus, Chlamydia and Mycoplasma, several viruses, and certain fungi and protozoans. The disease may be divided into two forms, bronchial pneumonia and lobar pneumonia. Bronchial pneumonia is most prevalent in infants, young children and aged adults. It is caused by various bacteria, including Streptococcus pneumoniae.Bronchial pneumonia involves the alveoli contiguous to the larger bronchioles of the bronchial tree. Lobar pneumonia is more prone to occur in younger adults. A majority (more than 80%) of the cases of lobar pneumonia are caused byStreptococcus pneumoniae. Lobar pneumonia involves all of a single lobe of the lungs (although more than one lobe may be involved), wherein the entire area of involvement tends to become a consolidated mass, in contrast to the spongy texture of normal lung tissue.
Bacteriology
Streptococcus pneumoniae cells are Gram-positive, lancet-shaped cocci (elongated cocci with a slightly pointed outer curvature). Usually, they are seen as pairs of cocci (diplococci), but they may also occur singly and in short chains. When cultured on blood agar, they are alpha hemolytic. Individual cells are between 0.5 and 1.25 micrometers in diameter. They do not form spores, and they are nonmotile. Like other streptococci, they lack catalase and ferment glucose to lactic acid. Unlike other streptococci, they do not display an M protein, they hydrolyze inulin, and their cell wall composition is characteristic both in terms of their peptidoglycan and their teichoic acid.
Gram Stain of a film of sputum from a case of lobar pneumonia. CDC.
Cultivation
Streptococcus pneumoniae is a fastidious bacterium, growing best in 5% carbon dioxide. Nearly 20% of fresh clinical isolates require fully anaerobic conditions. In all cases, growth requires a source of catalase (e.g. blood) to neutralize the large amount of hydrogen peroxide produced by the bacteria. In complex media containing blood, at 37°C, the bacterium has a doubling time of 20-30 minutes.
On agar, pneumococci grow as glistening colonies, about 1 mm in diameter. Two serotypes, types 3 and 37, are mucoid. Pneumococci spontaneously undergo a genetically determined, phase variation from opaque to transparent colonies at a rate of 1 in 105 . The transparent colony type is adapted to colonization of the nasopharynx, whereas the opaque variant is suited for survival in blood. The chemical basis for the difference in colony appearance is not known, but significant difference in surface protein expression between the two types has been shown.
Streptococcus pneumoniae is a fermentative aerotolerant anaerobe. It is usually cultured in media that contain blood. On blood agar, colonies characteristically produce a zone of alpha (green) hemolysis, which differentiates S. pneumoniaefrom the group A (beta hemolytic) streptococcus, but not from commensal alpha hemolytic (viridans) streptococci which are co-inhabitants of the upper respiratory tract. Special tests such as inulin fermentation, bile solubility, and optochin (an antibiotic) sensitivity must be routinely employed to differentiate the pneumococcus from Streptococcus viridans.
Streptococcus pneumoniae Gram-stain of blood broth culture. CDC.
Streptococcus pneumoniae is a very fragile bacterium and contains within itself the enzymatic ability to disrupt and to disintegrate the cells. The enzyme responsible is called an autolysin. The physiological role of this autolysin is to cause the culture to undergo a characteristic autolysis that kills the entire culture when grown to stationary phase. Virtually all clinical isolates of pneumococci harbor this autolysin and undergo lysis usually beginning between 18-24 hours after initiation of growth under optimal conditions. Autolysis is consistent with changes in colony morphology. Colonies initially appear with a plateau-type morphology, then start to collapse in the centers when autolysis begins.
Identification
The minimum criteria for identification and distinction of pneumococci from other streptococci are bile or optochin sensitivity, Gram-positive staining, and hemolytic activity. Pneumococci cause alpha hemolysis on agar containing horse, human, rabbit and sheep erythrocytes. Under anaerobic conditions they switch to beta hemolysis caused by an oxygen-labile hemolysin. Typically, pneumococci form a 16-mm zone of inhibition around a 5 mg optochin disc, and undergo lysis by bile salts (e.g. deoxycholate). Addition of a few drops of 10% deoxycholate at 37°C lyses the entire culture in minutes. The ability of deoxycholate to dissolve the cell wall, depends upon the presence of the autolytic enzyme, LytA. Virtually all clinical isolates of pneumococci harbor the autolysin and undergo deoxycholate lysis.
Streptococcus pneumoniae A mucoid strain on blood agar showing alpha hemolysis (green zone surrounding colonies). Note the zone of inhibition around a filter paper disc impregnated with optochin. Viridans streptococci are not inhibited by optochin.
Serotyping
The quellung reaction (swelling reaction) forms the basis of serotyping and relies on the swelling of the capsule upon binding of homologous antibody. The test consists of mixing a loopful of colony with equal quantity of specific antiserum and then examining microscopically at 1000X for capsular swelling. Although generally highly specific, cross-reactivity has been observed between capsular types 2 and 5, 3 and 8, 7 and 18, 13 and 30, and with E. coli, Klebsiella, H. influenzae Type b, and certain viridans streptococci.
Streptococcus pneumoniae Quellung (capsular swelling) reaction can be used to demonstrate the presence of a specific capsular type of the bacterium.
Cell Surface Structure
Streptococcus pneumoniae scanning electron micrograph of a pair of diplococci. CDC.
Capsule
A capsule composed of polysaccharide completely envelops the pneumococcal cells. During invasion the capsule is an essential determinant of virulence. The capsule interferes with phagocytosis by preventing complement C3b opsonization of the bacterial cells. 90 different capsule types of pneumococci have been identified and form the basis of antigenic serotyping of the organism. Anti-pneumococcal vaccines are based on formulations of various capsular (polysaccharide) antigens derived from the highly-prevalent strains.
A capsule composed of polysaccharide completely envelops the pneumococcal cells. During invasion the capsule is an essential determinant of virulence. The capsule interferes with phagocytosis by preventing complement C3b opsonization of the bacterial cells. 90 different capsule types of pneumococci have been identified and form the basis of antigenic serotyping of the organism. Anti-pneumococcal vaccines are based on formulations of various capsular (polysaccharide) antigens derived from the highly-prevalent strains.
Streptococcus pneumoniae Fluorescent antibody stain of capsular material. CDC.
Cell Wall
The cell wall of S. pneumoniae is roughly six layers thick and is composed of peptidoglycan with teichoic acid attached to approximately every third N-acetylmuramic acid. Lipoteichoic acid is chemically identical to the teichoic acid but is attached to the cell membrane by a lipid moiety. Both the teichoic acid and the lipoteichoic acid contain phosphorylcholine; two choline residues may be covalently added to each carbohydrate repeat. This is an essential element in the biology of S. pneumoniae since the choline specifically adheres to choline-binding receptors that are located on virtually all human cells.
The cell wall of S. pneumoniae is roughly six layers thick and is composed of peptidoglycan with teichoic acid attached to approximately every third N-acetylmuramic acid. Lipoteichoic acid is chemically identical to the teichoic acid but is attached to the cell membrane by a lipid moiety. Both the teichoic acid and the lipoteichoic acid contain phosphorylcholine; two choline residues may be covalently added to each carbohydrate repeat. This is an essential element in the biology of S. pneumoniae since the choline specifically adheres to choline-binding receptors that are located on virtually all human cells.
PiliHair-like structures that extend from the surface have recently been described in many strains of S. pneumoniae. They have been shown to contribute to colonization of the upper respiratory tract and to increase the formation of large amounts of TNF by the immune system during invasive infection.
Surface Proteins
On the basis of functional genomic analysis, it is estimated that the pneumococcus contains more than 500 surface proteins. Some are membrane-associated lipoproteins, and others are physically associated with the cell wall. The latter includes five penicillin binding proteins (PBPs), two neuraminidases, and an IgA protease. A unique group of proteins on the pneumococcal surface is the family of choline-binding proteins (CBPs). Twelve CBPs are noncovalently bound to the choline moiety of the cell wall and are used to "snap" various different functional elements onto the bacterial surface. The CBPs all share a common C-terminal choline-binding domain while the N-termini of the CBPs are distinct, indicating their functions are different. The CBP family includes such important determinants of virulence such as PspA (protective antigen), LytA, B, and C (three autolysins), and CbpA (an adhesin).
On the basis of functional genomic analysis, it is estimated that the pneumococcus contains more than 500 surface proteins. Some are membrane-associated lipoproteins, and others are physically associated with the cell wall. The latter includes five penicillin binding proteins (PBPs), two neuraminidases, and an IgA protease. A unique group of proteins on the pneumococcal surface is the family of choline-binding proteins (CBPs). Twelve CBPs are noncovalently bound to the choline moiety of the cell wall and are used to "snap" various different functional elements onto the bacterial surface. The CBPs all share a common C-terminal choline-binding domain while the N-termini of the CBPs are distinct, indicating their functions are different. The CBP family includes such important determinants of virulence such as PspA (protective antigen), LytA, B, and C (three autolysins), and CbpA (an adhesin).
Genetics
S. pneumoniae has a natural transformation system as a mechanism for genetic exchange. This process is of medical significance because it clearly underlies the explosion of antibiotic resistance in the bacterium over the past 20 years. For example, penicillin resistance is due to altered penicillin-binding proteins (PBPs) that exhibit a low affinity for beta lactam antibiotics. Comparison of the nucleotide sequences encoding the PBPs in S. pneumoniaeand S. mitis demonstrates that horizontal gene transfer has occurred between these two bacteria. In the laboratory, S. pneumoniae can also be transformed with genes from related and unrelated bacteria. As well, in the upper respiratory tract of the host, horizontal exchanges of genetic information could take place between strains of pneumococci that co-habitate or compete for dominance as normal flora.
Streptococcus pneumoniae can also develop antibiotic resistance by the timeless process of mutation and selection. The bacterium has a relatively fast growth rate and achieves large cell densities in an infectious setting, These conditions not only favor the occurrence natural transformation but also the emergence of spontaneous mutants resistant to the antibiotic.
During transformation, the binding, uptake and incorporation of exogenous DNA occur as a sequence of programmed events during a physiologically defined state known as competence. Competent bacteria self-aggregate, easily form protoplasts, are prone to autolysis and have an increased H+ and Na+ content that leads to increased glycolysis and enhanced ATP reserves. A unique set of at least 11 proteins is preferentially expressed during competence. Early in the competent state, a 17 amino acid peptide, known as competence-stimulating peptide (CSP) is released from the growing bacteria. CSP induces competence when it reaches a critical concentration that depends on the cell density, consistent with a quorum-sensing model.
athogenesis
Pneumococci spontaneously cause disease in humans, monkeys, rabbits, horses, mice and guinea pigs. Nasopharyngeal colonization occurs in approximately 40% of the population. Pneumonia and otitis media are the most common infections, meningitis being much more variable. The rabbit and the mouse have been used extensively as animal models of disease, leading to a reasonable understanding of many of the pneumococcal determinants of virulence.
Pneumococci spontaneously cause disease in humans, monkeys, rabbits, horses, mice and guinea pigs. Nasopharyngeal colonization occurs in approximately 40% of the population. Pneumonia and otitis media are the most common infections, meningitis being much more variable. The rabbit and the mouse have been used extensively as animal models of disease, leading to a reasonable understanding of many of the pneumococcal determinants of virulence.
Colonization
Pneumococci adhere tightly to the nasopharyngeal epithelium by multiple mechanisms that, for most individuals, appears to result in an immune response that generates type-specific immunity. For some people, however, progression into the lungs or middle ear occurs. Passage of pneumococci up the eustachian tube is accompanied by bacterial induced changes in the surface receptors of the epithelial cell, particularly by neuraminidase. Inflammation in the middle ear is caused by pneumococcal cell wall components, and pneumolysin inflicts major cytotoxicity on ciliated cells of the cochlea.
Pneumococci adhere tightly to the nasopharyngeal epithelium by multiple mechanisms that, for most individuals, appears to result in an immune response that generates type-specific immunity. For some people, however, progression into the lungs or middle ear occurs. Passage of pneumococci up the eustachian tube is accompanied by bacterial induced changes in the surface receptors of the epithelial cell, particularly by neuraminidase. Inflammation in the middle ear is caused by pneumococcal cell wall components, and pneumolysin inflicts major cytotoxicity on ciliated cells of the cochlea.
Upon reaching the lower respiratory tract by aerosol, pneumococci bypass the ciliated upper respiratory epithelial cells unless there is damage to the epithelium. Instead, they progress to the alveolus and associate with specific alveolar cells which produce a choline-containing surfactant.
Experimentally, in healthy tissues, it requires approximately 100,000 bacteria/ml to trigger an inflammatory response. However, if a proinflammatory signal is supplied, inflammation ensues with as few as 10 bacteria. This signal is a cytokine in experimental systems or an intercurrent viral infection in clinical situations. The inflammatory response can cause considerable tissue damage.
Invasion
The bacteria invade and grow primarily due to their resistance to the host phagocytic response. The cell wall components directly activate multiple inflammatory cascades, including the alternative pathway of complement activation, the coagulation cascade and the cytokine cascade, inducing interleukin-1, interleukin-6 and tumor necrosis factor (TNF) from macrophages and other cells.
The bacteria invade and grow primarily due to their resistance to the host phagocytic response. The cell wall components directly activate multiple inflammatory cascades, including the alternative pathway of complement activation, the coagulation cascade and the cytokine cascade, inducing interleukin-1, interleukin-6 and tumor necrosis factor (TNF) from macrophages and other cells.
In addition, as pneumococci begin to lyse due to autolysis or in response to host defensins and antimicrobial agents, they release cell wall components, pneumolysin and other substances that lead to greater inflammation and cytotoxic effects. Pneumolysin and hydrogen peroxide produced by the bacteria kill cells and induce production of nitric oxide which may play a key role in septic shock.
During invasion, the interaction between the bacterial cell wall choline and the host PAF receptor G-protein contributes to a state of altered vascular permeability. In the lung, this leads to arrival of an inflammatory exudate. At first, a serous exudate forms. This is followed by the arrival of leukocytes, thereby making the switch from a serous to a purulent exudate. Sites of pneumococcal infection are particularly noted for the intensity of the purulent response.
Pneumococci occasionally are able to directly invade endothelial cells. The ligands by which pneumococci bind to activated human cells include choline located on the cell wall teichoic acid that can serve as a direct ligand to the PAF receptor, and the choline-binding protein, CbpA, which binds to a specific carbohydrate on the alveolar cell surface. When bound to the PAF receptor, the pneumococcus enters a vacuole in a receptor-mediated endocytic process, and the vacuole moves across the cell expelling the bacteria on the ablumenal surface. In vitro, pneumococci will adhere to and traverse an endothelial barrier over approximately 4 hours.
If bacteremia occurs, the risk of meningitis increases. Pneumococci can adhere specifically to cerebral capillaries using the same pairings of choline to PAF receptor and CbpA to carbohydrate receptor. Thus, the bacteria subvert the endocytosis/recycling pathway of the PAF receptor for cellular transmigration. Once in the cerebrospinal fluid, a variety of pneumococcal components, particularly cell wall components, incite the inflammatory response.
Bacterial Determinants of Virulence
PiliThe initial event in invasive pneumococcal disease is the attachment of encapsulated pneumococci to epithelial cells in the upper respiratory tract. Recently, it has been shown that initial bacterial adhesion and subsequent ability to cause invasive disease is enhanced by pili, which were previously unknown to exist in pneumococci. These adhesive pili-like appendages are encoded by therlrA islet, present in some, but not all, clinical isolates. Introduction of the rlrA islet into an encapsulated rlrA-negative isolate allowed pilus expression, enhances adherence to lung epithelial cells, and provides a competitive advantage upon mixed intranasal challenge of mice. Furthermore, pilus-expressing rlrA islet-positive clinical isolates are more virulent than nonpiliated deletion mutants, and they out-compete the mutants in murine models of colonization, pneumonia, and bacteremia. Additionally, piliated pneumococci evoke a higher TNF response during systemic infection compared with nonpiliated derivatives, suggesting that pneumococcal pili not only contribute to adherence and virulence but also stimulate the host inflammatory response.
Capsule
The bacterial capsule interferes with phagocytosis by leukocytes, a property dependent on its chemical composition. Apparently, resistance to phagocytosis is brought about by interference with binding of complement C3b to the cell surface.
Capsule
The bacterial capsule interferes with phagocytosis by leukocytes, a property dependent on its chemical composition. Apparently, resistance to phagocytosis is brought about by interference with binding of complement C3b to the cell surface.
During invasion of the mucosal surface, encapsulated strains are 100,000 times more virulent than unencapsulated strains. The polysaccharide is nontoxic and noninflammatory, and the capsule does not appear to engage any host defenses except for the induction of antibody-mediated immunity. The pneumococcal capsule is not an antigenic disguise, and it does not impede the activities of underlying components, such as the cell wall and surface proteins, to engage the host defense systems. However, C-reactive protein or antibodies to teichoic acid, both of which bind to the cell wall under the capsule, fail to opsonize encapsulated strains.
Cell Wall Components
The pneumococcal cell wall is a collection of potent inflammatory stimuli. Challenge with cell wall components alone can recreate many of the symptoms of pneumonia, otitis media and meningitis in experimental models. The phosphorylcholine decorating the teichoic acid and the lipoteichoic acid is a key molecule enabling invasion, and acts both as an adhesin and as a docking site for the choline-binding proteins (CBPs). Other respiratory pathogens such asHaemophilus, Pseudomonas, Neisseria and Mycoplasma also have phosphorylcholine on lipopolysaccharide, proteins or pili, suggesting a shared mechanism for invasion of the respiratory tract. Two host-derived elements that recognize choline are platelet activating factor (PAF) receptor and the C-reactive protein. Since respiratory pathogens may be recognized and cleared by the C-reactive protein response as part of the innate defenses, respiratory pathogens may share this invasive mechanism to subvert the signaling cascade of endogenous PAF.
The pneumococcal cell wall is a collection of potent inflammatory stimuli. Challenge with cell wall components alone can recreate many of the symptoms of pneumonia, otitis media and meningitis in experimental models. The phosphorylcholine decorating the teichoic acid and the lipoteichoic acid is a key molecule enabling invasion, and acts both as an adhesin and as a docking site for the choline-binding proteins (CBPs). Other respiratory pathogens such asHaemophilus, Pseudomonas, Neisseria and Mycoplasma also have phosphorylcholine on lipopolysaccharide, proteins or pili, suggesting a shared mechanism for invasion of the respiratory tract. Two host-derived elements that recognize choline are platelet activating factor (PAF) receptor and the C-reactive protein. Since respiratory pathogens may be recognized and cleared by the C-reactive protein response as part of the innate defenses, respiratory pathogens may share this invasive mechanism to subvert the signaling cascade of endogenous PAF.
The peptidoglycan/teichoic acid complex of the pneumococcus is highly inflammatory. Smaller components of peptidoglycan progressively lose specific inflammatory activity. The cell wall directly activates the alternative pathway of the complement cascade, generating chemotaxins for leukocytes, and the coagulation cascade, which promotes a "procoagulant state" favoring thrombosis. In addition, peptidoglycan binds to CD14, a cell surface receptor known to initiate the inflammatory response for endotoxin. This induces a cytokine cascade resulting in production of interleukin-1, interleukin-6 and tumor necrosis factor from human cells.
Choline Binding Proteins
The CBP family includes such important determinants as PspA (protective antigen), LytA, B, and C (three autolysins), and CbpA (an adhesin).
The CBP family includes such important determinants as PspA (protective antigen), LytA, B, and C (three autolysins), and CbpA (an adhesin).
The protective antigen (PspA) is a 6 kDa protein with 10 choline-binding repeats. PspA appears to inhibit complement-mediated opsonization of pneumococci, and mutants lacking PspA have reduced virulence. Antibodies against PspA confer passive protection in mice.
Autolysin LytA is responsible for pneumococcal lysis in stationary phase as well as in the presence of antibiotics. The protein has two functional domains: a C-terminal domain with six choline-binding repeats that anchor the protein on the cell wall, and an N-terminal domain that provides amidase activity. Autolysin LytB is a glucosaminidase involved in cell separation, and LytC exhibits lysozyme-like activity.
CbpA is a major pneumococcal adhesin. It has eight choline-binding repeats. The adhesin interacts with carbohydrates on the pulmonary epithelial surface carbohydrates. CbpA-deficient mutants are defective in colonization of the nasopharynx and fail to bind to various human cells in vitro. CbpA also has been reported to bind secretory IgA and complement component C3.
Hemolysins
In addition to surface-associated virulence determinants, pneumococci secrete exotoxins. Two hemolysins have been described, the most potent of which is pneumolysin. Pneumolysin is a 53kDa protein that can cause lysis of host cells and activate complement. It is stored intracellularly and is released upon lysis of pneumococci. Pneumolysin binds to cholesterol and thus can indiscriminately bind to all cells without restriction to a receptor. The protein assembles into oligomers to form transmembrane pores which ultimately lead to cell lysis. Pneumolysin can also stimulate the production of inflammatory cytokines, inhibit beating of the epithelial cell cilia, inhibit lymphocyte proliferation, decrease the bactericidal activity of neutrophils, and activate complement. A second hemolysin activity has been described but has not been identified. In addition, pneumococci also produce hydrogen peroxide in amounts greater than human leukocytes produce. This small molecule is also a potent hemolysin.
In addition to surface-associated virulence determinants, pneumococci secrete exotoxins. Two hemolysins have been described, the most potent of which is pneumolysin. Pneumolysin is a 53kDa protein that can cause lysis of host cells and activate complement. It is stored intracellularly and is released upon lysis of pneumococci. Pneumolysin binds to cholesterol and thus can indiscriminately bind to all cells without restriction to a receptor. The protein assembles into oligomers to form transmembrane pores which ultimately lead to cell lysis. Pneumolysin can also stimulate the production of inflammatory cytokines, inhibit beating of the epithelial cell cilia, inhibit lymphocyte proliferation, decrease the bactericidal activity of neutrophils, and activate complement. A second hemolysin activity has been described but has not been identified. In addition, pneumococci also produce hydrogen peroxide in amounts greater than human leukocytes produce. This small molecule is also a potent hemolysin.
PiliAs mentioned above, pili contribute to colonization of upper respiratory tract and increase the formation of large amounts of tumor necrosis factor.
Hydrogen peroxideH2O2 produced by the pneumococcus causes damage to host cells (e.g. can cause apoptosis in neuronal cells during meningitis) and has bactericidal effects against competing bacteria such as Staphylococcus aureus.
Neuraminidase and IgA protease
These exoenzymes produced by the bacteria have a presumptive role in virulence as they do in other pathogens.
These exoenzymes produced by the bacteria have a presumptive role in virulence as they do in other pathogens.
Epidemiology
S. pneumoniae is a transient member of the normal flora, colonizing the nasopharynx of up to 40% of healthy adults and children with no adverse effects. Children carry this pathogen in the nasopharynx asymptomatically for about 4-6 weeks, often several serotypes at a time. New serotypes are acquired approximately every 2 months. Serotypes 6, 14, 18, 19, and 23 are the most prevalent, accounting for 60-80% of infections depending on the area of the world. Pneumococcal infection accounts for more deaths than any other vaccine-preventable bacterial disease. Those most commonly at risk for pneumococcal infection are children between 6 months and 4 years of age and adults over 60 years of age. Virtually every child will experience pneumococcal otitis media before the age of 5 years. It is estimated that 25% of all community-acquired pneumonia is due to pneumococcus (1,000 per 100,000 inhabitants).
Until 2000, S. pneumoniae infections caused 100,000-135,000 hospitalizations for pneumonia, 6 million cases of otitis media, and 60,000 cases of invasive disease, including 3300 cases of meningitis. (CDC reported 60,000 cases of invasive pneumococcal disease in 1997, resulting in approximately 6,000 deaths.) The incidence of sterile-site infections haS shown geographic variation from 21 to 33 cases per 100,000 population. Disease figures are now changing due to conjugate vaccine introduction. In 2002, the rate of invasive disease was 13 cases per 100,000 in the United States. However, epidemics of disease have reappeared in settings such as chronic care facilities, military camps and day care centers, a situation not recognized since the pre-antibiotic era.
Also of concern, is the increased emergence of antibiotic resistance, especially in the past two decades. Multiple antibiotic resistant strains of S. pneumoniae that emerged in the early 1970s in Papua New Guinea and South Africa were thought to be a fluke, but multiple antibiotic resistance now covers the globe and has rapidly increased since 1995. Increases in penicillin resistance have been followed by resistance to cephalosporins and multidrug resistance. The incidence of resistance to penicillin increased from <0.02 in 1987 to 3% in 1994 to 30% in some communities in the United States and 80% in regions of some other countries in 1998. Resistance to other antibiotics has emerged simultaneously: 26% resistant to trimethoprim-sulfa, 9% resistant to cefotaxime, 30% resistant to macrolides, and 25% resistant to multiple drugs. Resistant organisms remain fully virulent but seem to have arisen in less than 10 serotypes. Serotypes 6A, 6B, 9V, 14, 19A and 23F are included in the vast majority of resistant strains.
Vaccines
Given the 90 different capsular types of pneumococci, a comprehensive vaccine based on polysaccharide alone is not yet feasible. Thus, vaccines based on a subgroup of highly prevalent types have been formulated. The number of serotypes in the vaccine has increased from four in 1945, to 14 in the 1970s, and finally to the current 23-valent formulation (25 mg of each of serotypes 1, 2, 3, 4, 5, 6B, 7F, 8, 9N, 9V, 10A, 11A, 12F, 14, 15B, 17F, 18C, 19A, 19F, 20, 22F, 23F, and 33F). These serotypes represent 85-90% of those that cause invasive disease, and the vaccine efficacy is estimated at 60% . However, underutilization of the vaccine is so extensive that the pneumococcus remains the most common infectious agent leading to hospitalization in all age groups. This is further complicated by the fact that polysaccharides are not immunogenic in children under the age of 2 years where a significant amount of disease occurs. Immunization is suggested for those at highest risk of infection, including those 65 years or older, and generally should be a single lifetime dose.
In the United States, a heptavalent pneumococcal conjugate vaccine (PCV7) has been recommended since 2000 for all children aged 2-23 months and for at-risk children aged 24-59 months. The four-dose series is given at 2, 4, 6 and 12-14 months of age. Protection is good against against invasive pneumococcal infections, especially septicemia and meningitis. However, children exposed to a serotype not contained in the vaccine are not afforded any protection. This limitation, and the ability of capsular-polysaccharide conjugate vaccines to promote the spread of non-covered serotypes, has led to research into vaccines that would provide species-wide protection.
No comments:
Post a Comment