A Southern blot is a method used in molecular biology for detection of a specific DNA sequence in DNA samples. Southern blotting combines transfer of electrophoresis-separated DNA fragments to a filter membrane and subsequent fragment detection by probe hybridization.
The method is named after its inventor, the British biologist Edwin Southern. Other blotting methods (i.e., western blot, northern blot, eastern blot, southwestern blot) that employ similar principles, but using RNA or protein, have later been named in reference to Edwin Southern's name. As the technique was eponymously named, Southern blot is capitalized as is conventional for proper nouns. The names for other blotting methods may follow this convention, by analogy
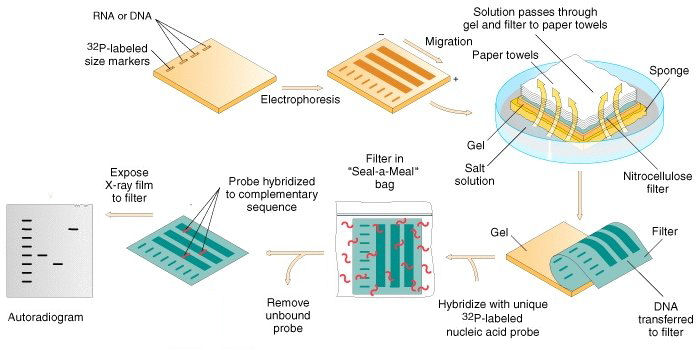
Method
Restriction endonucleases are used to cut high-molecular-weight DNA strands into smaller fragments.
The DNA fragments are then electrophoresed on an agarose gel to separate them by size.
If some of the DNA fragments are larger than 15 kb, then prior to blotting, the gel may be treated with an acid, such as dilute HCl. This depurinates the DNA fragments, breaking the DNA into smaller pieces, thus allowing more efficient transfer from the gel to membrane.
If alkaline transfer methods are used, the DNA gel is placed into an alkaline solution (typically containing sodium hydroxide) to denature the double-stranded DNA. The denaturation in an alkaline environment may improve binding of the negatively charged thymine residues of DNA to a positively charged amino groups of membrane, separating it into single DNA strands for later hybridization to the probe (see below), and destroys any residual RNA that may still be present in the DNA. The choice of alkaline over neutral transfer methods, however, is often empirical and may result in equivalent results.
A sheet of nitrocellulose (or, alternatively, nylon) membrane is placed on top of (or below, depending on the direction of the transfer) the gel. Pressure is applied evenly to the gel (either using suction, or by placing a stack of paper towels and a weight on top of the membrane and gel), to ensure good and even contact between gel and membrane. If transferring by suction 20X SSC buffer is used to ensure a seal and prevent drying of the gel. Buffer transfer by capillary action from a region of high water potential to a region of low water potential (usually filter paper and paper tissues) is then used to move the DNA from the gel on to the membrane; ion exchange interactions bind the DNA to the membrane due to the negative charge of the DNA and positive charge of the membrane.
The membrane is then baked in a vacuum or regular oven at 80 °C for 2 hours (standard conditions; nitrocellulose or nylon membrane) or exposed to ultraviolet radiation (nylon membrane) to permanently attach the transferred DNA to the membrane.
The membrane is then exposed to a hybridization probe—a single DNA fragment with a specific sequence whose presence in the target DNA is to be determined. The probe DNA is labelled so that it can be detected, usually by incorporating radioactivity or tagging the molecule with a fluorescent or chromogenic dye. In some cases, the hybridization probe may be made from RNA, rather than DNA. To ensure the specificity of the binding of the probe to the sample DNA, most common hybridization methods use salmon or herring sperm DNA for blocking of the membrane surface and target DNA, deionized formamide, and detergents such as SDS to reduce non-specific binding of the probe.
After hybridization, excess probe is washed from the membrane (typically using SSC buffer), and the pattern of hybridization is visualized on X-ray film by autoradiography in the case of a radioactive or fluorescent probe, or by development of color on the membrane if a chromogenic detection method is used.
Result
Hybridization of the probe to a specific DNA fragment on the filter membrane indicates that this fragment contains DNA sequence that is complementary to the probe. The transfer step of the DNA from the electrophoresis gel to a membrane permits easy binding of the labeled hybridization probe to the size-fractionated DNA. It also allows for the fixation of the target-probe hybrids, required for analysis by autoradiography or other detection methods. Southern blots performed with restriction enzyme-digested genomic DNA may be used to determine the number of sequences (e.g., gene copies) in a genome. A probe that hybridizes only to a single DNA segment that has not been cut by the restriction enzyme will produce a single band on a Southern blot, whereas multiple bands will likely be observed when the probe hybridizes to several highly similar sequences (e.g., those that may be the result of sequence duplication). Modification of the hybridization conditions (for example, increasing the hybridization temperature or decreasing salt concentration) may be used to increase specificity and decrease hybridization of the probe to sequences that are less than 100% similar.
Applications
Southern transfer may be used for homology-based cloning on the basis of amino acid sequence of the protein product of the target gene. Oligonucleotides are designed that are similar to the target sequence. The oligonucleotides are chemically synthesised, radiolabeled, and used to screen a DNA library, or other collections of cloned DNA fragments. Sequences that hybridise with the hybridisation probe are further analysed, for example, to obtain the full length sequence of the targeted gene. Second, Southern blotting can also be used to identify methylated sites in particular genes. Particularly useful are the restriction nucleases MspI and HpaII, both of which recognize and cleave within the same sequence. However, HpaII requires that a C within that site be methylated, whereas MspI cleaves only DNA unmethylated at that site. Therefore, any methylated sites within a sequence analyzed with a particular probe will be cleaved by the former, but not the latter, enzyme.
No comments:
Post a Comment